|
|
撰写:花博 来源:小张聊科研平台的“ i生信”公众号,微信公众号搜索“ i生信”即可关注/扫描关注见文末
#科研热点# #代谢重编程#
大家好,又到了不定期更新“细胞表型怎么选”系列推文的时间,继上次上上次上上上次为大家介绍了细胞程序性死亡(凋亡坏死焦亡铁死亡不知道怎么选?看完这篇从此不再纠结)、细胞衰老(这个人人回避的问题,竟是科研热点?)、铜代谢(【热点追踪】铜代谢到底是个啥?我们这就帮你整理好了)之后,这回要给大家隆重推出的国自然研究热点表型是——代谢重编程。
有人看了之后会不屑一顾地说,切~不就是那个什么瓦什么格效应吗,早就知道啦(Otto Heinrich Warburg:我没有惹你们任何人)。的确,Warburg效应,更通俗的说法为肿瘤的有氧糖酵解,是肿瘤代谢重编程中非常重要的一种机制,Warburg在20世纪初提出这一假说后不断被后来的研究证实,距今已经历了近百年。

(图片来源于网络)O. H. Warburg这张模糊的黑白照甚至有点神似憨豆先生
然而,随着对肿瘤代谢研究的逐渐深入,如今代谢重编程早已不只是Warburg效应的代名词,而是广义上指一切细胞代谢机制改变的过程。那么代谢重编程到底包含哪些研究方向,又怎样结合临床问题设计课题呢?且看花博为大家一一解答。
1.什么是代谢重编程?
从广义上说,细胞代谢重编程是指细胞为满足能量需求,通过改变代谢模式促进细胞存活和生长的机制,其不仅能帮助细胞抵御外界胁迫,还能赋予细胞新的功能。因此,参与细胞能量代谢的基本物质都有可能发生代谢重编程。
我们最熟悉的Warburg效应,就是肿瘤细胞为了自身能量需要,将主要的葡萄糖代谢方式从氧化磷酸化(OXPHO)转变为糖酵解(glycolysis)的过程。除此之外,目前发现的代谢重编程还包括脂肪酸、氨基酸代谢的重编程。
1)葡萄糖代谢重编程(有氧糖酵解)
正常细胞在常氧条件下通过呼吸作用,即氧化磷酸化(OXPHO)产生ATP,在低氧/无氧条件下才通过糖酵解产能。然而一些肿瘤细胞为了满足对能量的需求代谢方式发生改变,即使在常氧情况下也通过糖酵解产生ATP,这一过程称为有氧糖酵解,即Warburg效应。需要注意的是,肿瘤细胞并不是在所有阶段、所有状态下都处于有氧糖酵解占主导,在不同癌症类型、发展阶段和外部干预下,肿瘤细胞会根据情况转换有氧糖酵解和OXPHO。

肿瘤发展过程中的代谢进化[1]
2)脂肪酸代谢重编程
提到脂肪酸代谢大家可能第一反应想到的是脂肪细胞,但其实在其它细胞中脂肪酸代谢同样是能量代谢过程中不可缺少的一环。尤其对于肿瘤来说,脂肪酸代谢重编程影响肿瘤的发生发展、侵袭转移,脂肪酸代谢产物甚至可作为临床诊断标志物,与患者不良预后有关[2]。
脂肪酸氧化(FAO)产生乙酰辅酶A(Acetyl-CoA)参与三羧酸(TCA)循环,一旦FAO发生异常或障碍,将直接影响细胞能量生产模式。
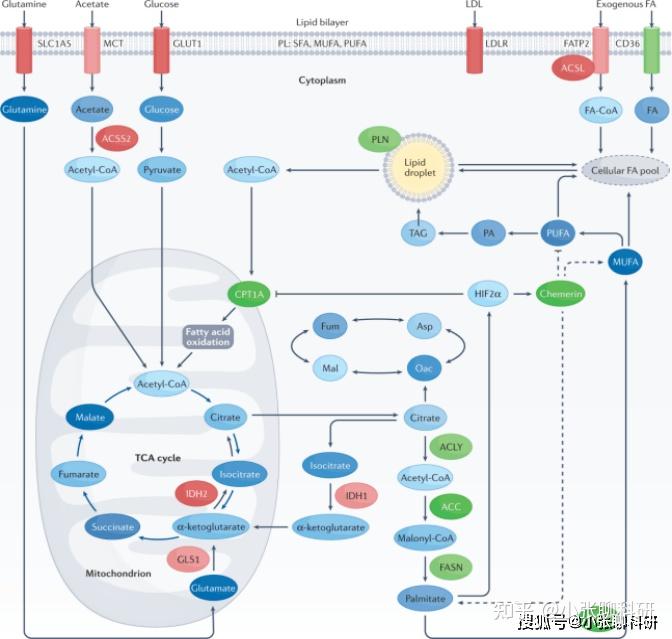
肾透明细胞癌脂肪酸代谢途径和潜在靶点[2]
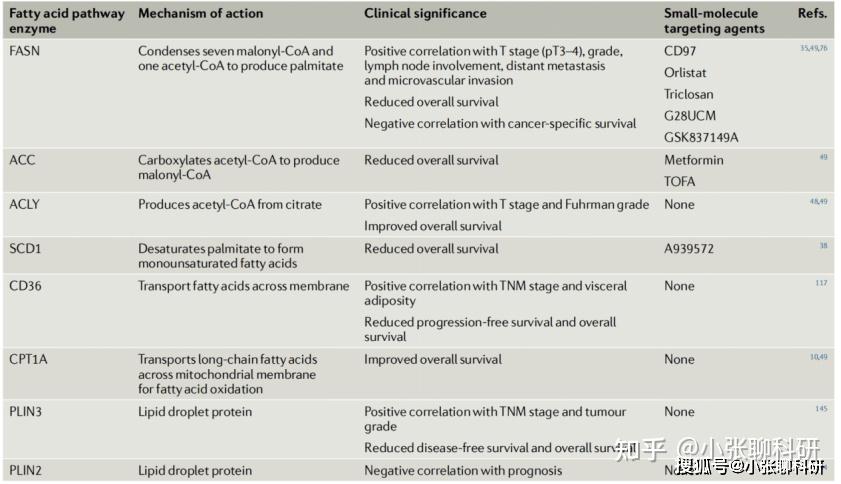
而与脂肪酸代谢相关的酶,如脂肪酸合成酶、乙酰辅酶A羧化酶、脂肪酸转运酶等,均可能参与到脂肪酸代谢重编程和肿瘤行为,部分酶已被作为治疗靶点设计小分子靶向药物,如下图所示。
3)氨基酸代谢重编程
氨基酸代谢重编程,类比前面介绍的糖代谢和脂肪酸代谢,就是细胞改变某种或某些氨基酸的代谢模式以满足自身活动需要。在肿瘤中,氨基酸代谢的改变显著影响微环境和肿瘤免疫,进而与免疫逃逸、耐药相关。
由于免疫细胞,尤其是淋巴细胞对氨基酸浓度十分敏感,因此氨基酸代谢重编程对肿瘤浸润的免疫细胞影响最大。已发现T细胞表面的氨基酸转运体主要为SLC家族蛋白,受到T细胞受体信号通路调控,如下图示。
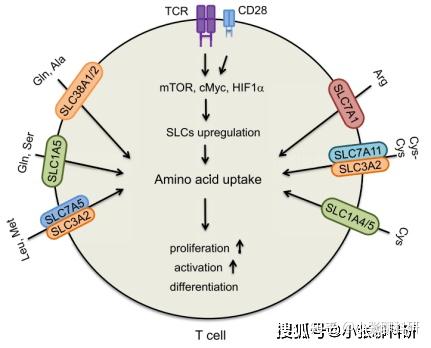
T细胞表面的氨基酸转运体及其功能[3]
除了T细胞之外,肿瘤细胞表面同样表达氨基酸转运体,因此T细胞的氨基酸代谢重编程又会反向作用于肿瘤,影响肿瘤细胞行为。比如甲硫氨酸在肿瘤细胞内促进蛋白质的合成,或转化为组蛋白甲基化的供体分子SAM。
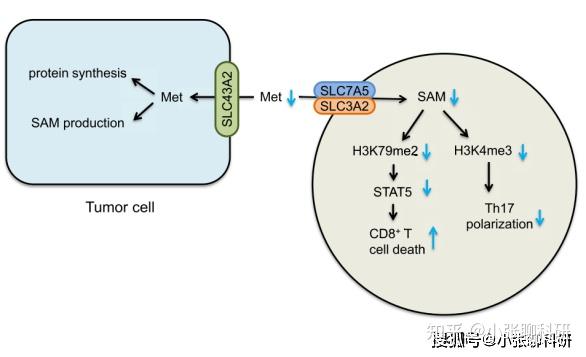
甲硫氨酸水平对肿瘤细胞和T细胞功能的调控作用[3]
2.代谢重编程发生在哪里?诱导因素有哪些?
从细胞定位角度考虑,代谢重编程主要发生在线粒体和细胞质中;从细胞类型角度考虑,无论是肿瘤细胞还是非肿瘤细胞,比如免疫细胞、成纤维细胞、上皮细胞,甚至是病原体都有可能发生代谢重编程。
代谢重编程归根到底涉及代谢相关蛋白的表达差异和酶活变化,因此依然与细胞内的调控及响应机制息息相关。以肿瘤举例,从内因上来说,促癌基因型、表观遗传模式、扰动的信号和基因表达、停滞的发育状态都是代谢重编程的触发因素;而外因则更广泛,包括组织结构、微环境内其它细胞的行为、细胞间互作、异常的代谢环境、肥胖/糖尿病等代谢疾病,这些都可能诱导肿瘤代谢重编程。
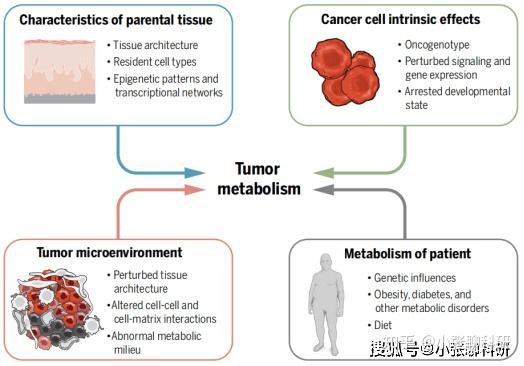
影响肿瘤代谢的内因与外因[1]
3.与代谢重编程相关的疾病有哪些?
除了肿瘤以外,在非肿瘤疾病领域也存在许多代谢重编程的证据。比如作为葡萄糖消耗量最大的器官——大脑,就会发生有氧糖酵解,虽然大脑是一个高度耗氧的器官,但它在激活过程中优先上调葡萄糖的糖酵解代谢。有氧糖酵解在整体和局部水平都发生,包括三个主要途径:产生和释放乳酸的糖酵解、磷酸戊糖分流和星形胶质细胞的糖原转化。已知有氧糖酵解是由肾上腺素能调节,但具体机制仍不清楚。因此,与神经系统相关的疾病与代谢重编程关系密切。
此外,如前文所述,目前对免疫细胞的代谢重编程关注度最高,涉及的细胞类型包括巨噬细胞、T细胞、B细胞、NK细胞等,包括肿瘤疾病和一些有免疫细胞参与的疾病。
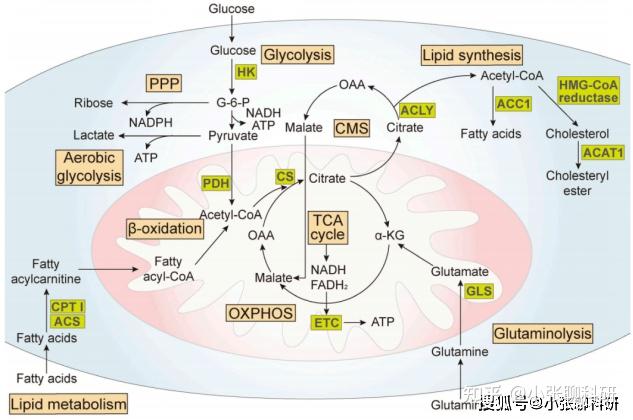
代谢重编程涉及的主要通路[4]
除此之外,病理状态下的普通组织细胞类型,如上皮细胞也可能发生代谢重编程,下面这篇今年发表在《Cell Reports》上的文章研究的就是干眼症中角膜上皮细胞,由于暴露于轻度渗透压力,诱导细胞内氨基酸转运蛋白SNAT2导致代谢重编程的分子机制[5]。

(doi: 10.1016/j.celrep.2022.111092)
以下为大家展示部分代谢重编程国自然中标标题,说不定可以打开思路,获得灵感。

4.代谢重编程的课题设计思路
最后再来讲讲代谢重编程的课题设计。与其它可观察表型的热点有所不同,代谢重编程并不是一个可以直接观察就发现的表型。因此通常我们都是通过代谢组学或者检测指定代谢物的方式找到切入角度,再进一步筛选基因。
基本步骤是首先在数据中发现某种代谢物含量显著变化,然后通过转录组或蛋白组发现与该代谢物合成/降解直接相关的基因/酶(主角分子),将样本分为主角分子低表达和高表达组,比较两组之间疾病进展,或临床问题相关的病征、指标与主角分子表达是否相关。之后就是构建基因干预模型,确认代谢表型基础上,还需检测疾病相关指标表型(比如增殖侵袭迁移),进而筛选下游通路。最后就仍然是分子机制验证实验。
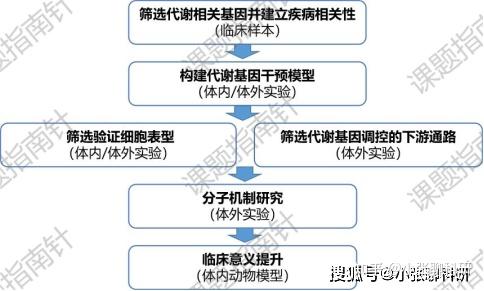
从创新性角度考虑,非肿瘤细胞类型的新颖性大于肿瘤细胞,氨基酸代谢重编程新颖程度大于脂代谢重编程,创新性最低的是糖代谢重编程(有氧糖酵解),大家可根据自己的实际情况选择。
如果想要看代谢重编程相关的研究思路和文献解读,还是要记得在下方留言哦!
参考文献:
[1] Faubert, B., Solmonson, A., & DeBerardinis, R. J. (2020). Metabolic reprogramming and cancer progression. Science (New York, N.Y.), 368(6487), eaaw5473.
[2] Tan, S. K., Hougen, H. Y., Merchan, J. R., Gonzalgo, M. L., & Welford, S. M. (2022). Fatty acid metabolism reprogramming in ccRCC: mechanisms and potential targets. Nature reviews. Urology, 10.1038.
[3] Wang, W., & Zou, W. (2020). Amino Acids and Their Transporters in T Cell Immunity and Cancer Therapy. Molecular cell, 80(3), 384–395.
[4] Yu, H., Jacquelot, N., & Belz, G. T. (2022). Metabolic features of innate lymphoid cells. The Journal of experimental medicine, 219(11), e20221140.
[5] Krokowski, D., Jobava, R., Szkop, K. J., Chen, C. W., Fu, X., Venus, S., Guan, B. J., Wu, J., Gao, Z., Banaszuk, W., Tchorzewski, M., Mu, T., Ropelewski, P., Merrick, W. C., Mao, Y., Sevval, A. I., Miranda, H., Qian, S. B., Manifava, M., Ktistakis, N. T., … Hatzoglou, M. (2022). Stress-induced perturbations in intracellular amino acids reprogram mRNA translation in osmoadaptation independently of the ISR. Cell reports, 40(3), 111092.
原文链接: |
|



 发表于 2022-12-21 19:47:37
发表于 2022-12-21 19:47:37